Развитие биологического ответа
Требуемые условия завершения
Самая первая из предложенных теорий принадлежала Кларку (1926), который предположил, что выраженность наблюдаемого эффекта связана с количеством рецепторов, оккупированных токсикантом:
ρ = f(Z).
Чаще всего эта зависимость считается линейной, т. е. реакция пропорциональна количеству занятых рецепторов (справедливо в случае малых концентраций эффектора).
В соответствии с рассматриваемой теорией токсикант может быть либо агонистом, либо антагонистом. Однако результаты многочисленных исследований указывают на то, что подобная классификация веществ оказывается недостаточной для описания наблюдаемых эффектов. Так установлено, что максимальный эффект, вызываемый различными агонистами, действующими на одну и туже рецепторную систему, не одинаков. Для преодоления этого противоречия Стефенсоном (1956) были предложены три допущения:
– максимальный эффект может быть вызван агонистом даже в том случае, если оккупирована лишь незначительная часть рецепторов;
– развивающийся эффект не линейно связан с количеством оккупированных рецепторов;
– токсиканты обладают неодинаковой эффективностью (относительной возбуждающей активностью), т.е. способностью вызывать эффект, взаимодействуя с рецептором.
В приведенных моделях предполагается существование неветвящейся последовательности реакций, первая из которых индуцируется образованием комплекса рецептор – эффектор.
В действительности же каждое звено развития индуцированного процесса может служить источником стимула для нескольких элементов. В результате, наблюдая за развитием реакции, мы регистрируем при малых концентрациях лишь сигнал, распространяющийся через наиболее чувствительный канал. По мере увеличения примененной концентрации проявляются эффекты иных каналов связи, сигналы которых, очевидно, могут отличаться по знаку (вклад отдельных сигналов может быть как положительным (стимулирующим реакцию), так и отрицательным (угнетающим).
Здесь можно рассмотреть два случая: 1) лиганд взаимодействует с мембранактивными центрами одного типа, а дифференциация характера действия происходит на каких-то следующих стадиях; 2) лиганд взаимодействует с центрами разных типов, каждый из которых инициирует процесс, различающийся по конечному эффекту в отношении регистрируемой реакции.
В первом случае справедливо представить реакцию в виде ρ = f(Z).
Второй случай – взаимодействие вещества с несколькими типами рецепторов (взаимодействие может происходить не только со специфическими рецепторами, но и с другими центрами связывания). Известно, например, что некоторые биологически активные пептиды уже в довольно низких концентрациях способны действовать неспецифически, т. е. помимо рецепторов модифицировать свойства клеточных мембран. Это несомненно должно отразиться и на развитии регистрируемой реакции.
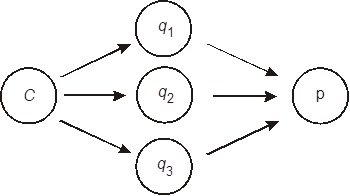
Рисунок 13 – Схема формирования реакции р под влиянием эффектора С, распространяющегося по нескольким путям. Каналы qt могут характеризоваться существенно различающимися константами сродства к эффектору
Последнее изменение: понедельник, 19 февраля 2018, 12:55